Back to Journals » Breast Cancer: Targets and Therapy » Volume 16
Intermediate Filaments in Breast Cancer Progression, and Potential Biomarker for Cancer Therapy: A Narrative Review
Authors Shalannandia WA, Chou Y ![]() , Bashari MH
, Bashari MH ![]() , Khairani AF
, Khairani AF ![]()
Received 3 August 2024
Accepted for publication 7 October 2024
Published 14 October 2024 Volume 2024:16 Pages 689—704
DOI https://doi.org/10.2147/BCTT.S489953
Checked for plagiarism Yes
Review by Single anonymous peer review
Peer reviewer comments 2
Editor who approved publication: Professor Harikrishna Nakshatri
Widad Aghnia Shalannandia,1 Yoan Chou,2 Muhammad Hasan Bashari,1,3 Astrid Feinisa Khairani1,3
1Graduate School of Master Program in Biomedical Sciences, Faculty of Medicine, Universitas Padjadjaran, Sumedang, Jatinangor, West Java, Indonesia; 2Graduate School of Master Program in Anti Aging and Aesthetic Medicine, Faculty of Medicine, Universitas Padjadjaran, Sumedang, Jatinangor, West Java, Indonesia; 3Department of Biomedical Sciences, Faculty of Medicine, Universitas Padjadjaran, Sumedang, Jatinangor, West Java, Indonesia
Correspondence: Astrid Feinisa Khairani, Department of Biomedical Sciences, Faculty of Medicine, Universitas Padjadjaran, Jalan Raya Bandung – Sumedang Km 21, Jatinangor, Sumedang, West Java, 45363, Indonesia, Tel +62-22-7795594, Email [email protected]
Abstract: Intermediate filaments are one of the three components of the cytoskeletons, along with actin and microtubules. The intermediate filaments consist of extensive variations of structurally related proteins with specific expression patterns in cell types. The expression pattern alteration of intermediate filaments is frequently correlated with cancer progression, specifically with the epithelial-to-mesenchymal transition process closely related to increasing cellular migration and invasion. This review will discuss the involvement of cytoplasmic intermediate filaments, specifically vimentin, nestin, and cytokeratin (CK5/CK6, CK7, CK8/CK18, CK17, CK19, CK20, CSK1), in breast cancer progression and as prognostic or diagnostic biomarkers. The potential for drug development targeting intermediate filaments in cancer will be reviewed.
Keywords: breast malignancy, cytokeratin, EMT, intermediate filament, keratin, nestin, vimentin
Graphical Abstract:
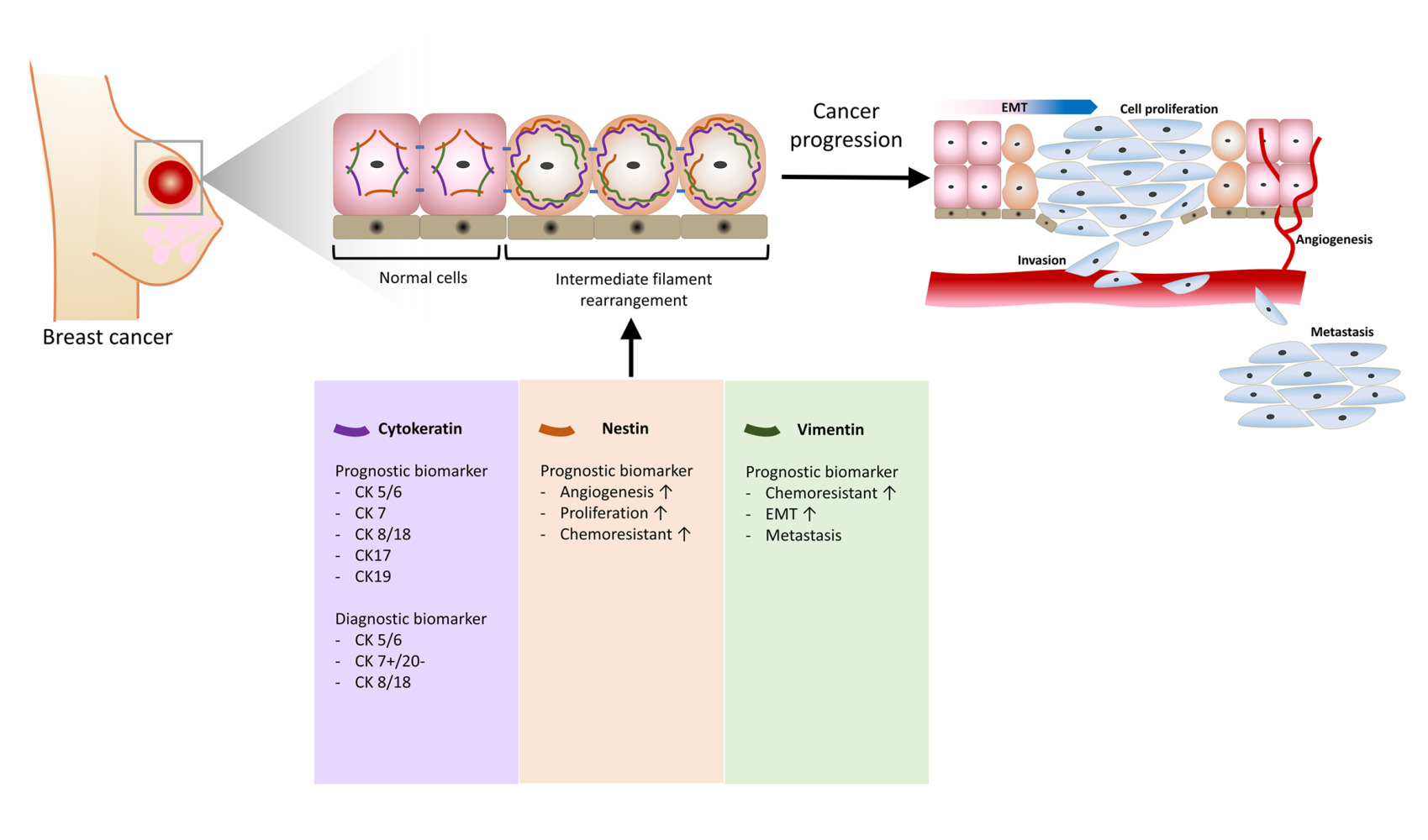
Introduction
During tumor progression, the mutated cells undergo malignant transformation that induces a sequence of structural rearrangements. This cellular transformation is closely related to the Epithelial-to-Mesenchymal Transition (EMT) process that allows the cells to gain mobility and leads to the ability of the cells to migrate and invade the blood vessel, leading to cancer metastasis at a distant site. Multiple biochemical modifications happen to epithelial cells that undergo the EMT process to gain mobility and extracellular matrix (ECM) component productibility of mesenchymal cells. The biochemical mechanism includes initiation of EMT by transcription factors, expression of specific cell-surface proteins, cytoskeletal modification and rearrangement, enzyme secretion for ECM degradation, and alteration in specific microRNA expression. Thus, it leads to cancer progression and metastasis to distant sites.1–5
The cytoskeleton’s intermediate filament plays a prominent role in maintaining cell integrity. The rearrangement of intermediate filaments, including vimentin, nestin, and keratins, has also been reported to be involved in cancer malignancy.6 Over the years, various intermediate filaments have been used in scientific and clinical studies as biomarkers to assist in identifying tumor types and prognoses and determining appropriate cancer treatment.7–9 The exploration of intermediate filaments has been studied in various cancers, including breast cancer. This cancer is ranked second after lung cancer for the mortality-to-incidence rate, which accounts for 15%.10 Breast cancer holds the first place globally as the highest cancer incidence in both sexes (11.7%). While in the female population, the incidence rate of breast cancer reaches 24.5%, with a mortality rate of 15.5%.11
This review will summarize the roles of intermediate filaments in cancer studies and the progression of breast cancer, focusing on three types of intermediate filaments: vimentin, nestin, and keratin. We will also discuss the potential of intermediate filaments as molecular targets for cancer therapy.
Intermediate Filament in Cancer Studies
The intermediate filament comprises the largest and the most diverse cytoskeletal protein family besides actin filament and microtubules.12 The intermediate filament is divided into six types (type I–VI). The first four types of intermediate filament (type I–IV) can be found in the cytoplasm of various cells. Type V of intermediate filament consists of lamins located in the nucleus, known as nuclear lamins. The last type of intermediate filament (type VI) can be found in the lens.6 The major characteristics of intermediate filaments are their rope-like structure and excellent strength, which help cells withstand mechanical stress and maintain their structural integrity. The intermediate filament is abundantly found in the cytoplasm of cells prone to mechanical shears, such as neural cells, skin epithelial cells, and mobile mesenchymal cells.13
In cancer studies, staging is generally determined by evaluating cancer size and its spreading to nearby tissue (T), lymph nodes (N), and distant organs or metastasis (M). Based on this TNM system, cancer progression can be classified into 5 stages (Table 1). Cancer development starts with the accumulation of abnormal changes or dysplasia in a group of cells called carcinoma in situ (CIS).14 CIS has the potential to develop into cancer later and form a primary tumor. The growth of primary tumors induces angiogenesis to assist in supplying nutrients to the tumor mass. The vasculature formed during angiogenesis facilitates the spreading of cancer cells or metastasis.15 However, the cells required an invasive capability to breach the endothelial layer and survive in the vascular system. Cancer cells can acquire this invasive property through the Epithelial-to-Mesenchymal transition (EMT) process. As the name mentioned, this process goal is acquiring the mesenchymal characteristics of the cells, in which the mobility and stress resistance of cancer cells increase. Hence, the cancer cells could survive until they reached the metastatic sites.1,16
 |
Table 1 Summary of TNM Cancer Staging System |
Substantial amounts of biomarkers, including various intermediate filaments, have been studied to determine the progression and prognosis of cancer.8,17,18 The intermediate filament property of cancer has been used as a prognostic biomarker to assist in the determination of an appropriate therapeutic regimen for breast cancer. In addition, some types of intermediate filament have also been reported to be associated with the cancer’s chemoresistance to particular chemotherapy regiments.19,20 Besides their role as prognostic biomarkers, the intermediate filaments have also been employed as diagnostic biomarkers and help characterize the specific type of breast cancer.21–23 In particular, the desmin intermediate filament has been utilized as a diagnostic marker for sarcomas, including breast sarcoma, a rare type of breast cancer. Desmin is a type III intermediate filament found in muscle and endothelial cells. Furthermore, it was used as one of the biomarkers to identify Rhabdomyosarcoma in male breast cancer patients in one of the case studies.24,25 Nevertheless, other intermediate filaments considerably explored in cancer studies include vimentin, nestin, and keratin, which will be discussed further in the following sections.
Vimentin
Vimentin is categorized as type III intermediate filaments. The vimentin and vimentin-related filaments are commonly found in connective tissue cells, muscle cells, and glial cells.6 As an intermediate filament, vimentin is crucial in cell extension formation, maintains cell integrity, and strengthens cells against mechanical stress. Vimentins also surround and protect the nucleus against deformation during a persistent invasion.26–28 With its various roles in diverse types of cells and tissue function, this protein is also closely related to many human diseases, including Crohn’s disease, HIV, cataracts, and cancer.29
In the cancer field, vimentin is often mentioned to be involved in the tumor cells’ epithelial-to-mesenchymal transition (EMT) process. Vimentin is abundantly expressed in mesenchymal cells while lowly expressed in epithelial cells. Thus, it is widely known as one of the markers of EMT.30 Furthermore, vimentin is closely involved in cancer metastasis, including migration, invasion, and adhesion regulation initiated by the EMT process. Vimentin promotes cell migration by inhibiting focal adhesion-associated proteins. Meanwhile, in contrast, the focal adhesion molecules will inhibit migration in the lack of vimentin expression.31,32 Moreover, another in vitro study on breast cancer reported that vimentin expression increased in the persistent cells upon chemotherapy. These findings indicated that the persistent cells had increased their invasiveness. Furthermore, the same study also reported that down-regulation of vimentin could reduce invasive capability while increasing sphere formation ability in MDA-MB-231 persistent cells. This outcome confirms that vimentin positively regulates the invasive capability of cancer cells.33
Various molecular pathways involve vimentin expression. One of the often-discussed pathways related to vimentin is the PI3K/AKT pathway, which also regulates the activation of EMT transcription factors.34,35 Earlier studies evaluated AKT activity in cell migration and its relation with vimentin as the direct substrate of AKT. The study demonstrated phosphorylation and activation of vimentin by AKT, specifically the AKT1. The AKT1-vimentin interaction mediates the induction of motility and invasion of tumor cells. Furthermore, the inhibition of AKT resulted in the activation of caspase 3 and vimentin proteolysis (Figure 1).34
 |
Figure 1 Vimentin involvement in cancer. |
MMPs family that has been reported to increase in various cancer cells are also involved in cancer progression and metastasis.36,37 MMPs activity is also stated to be involved in activating EMT through the down-regulation of epithelial adhesion molecules and upregulation of the mesenchymal features in the cells. The study by Li et al discovered that the inhibition of membrane-type 1 matrix metalloproteinase (MT1-MMP) attenuates vimentin expression in cancer cells. Moreover, this condition also suppresses the invasive capability of cancer cells. Hence, it suggests the crucial role of this MMPs family in EMT through the regulation of Vimentin (Figure 1).37
Furthermore, intriguing emerging exploration of the role of cell-surface vimentin (CSV) has been increasing in recent studies as a potential receptor for the development of cancer-specific targeting agents like monoclonal antibodies.38,39 Previous studies reported the potential role of CSV as a prognostic biomarker in detecting circulating tumor cells (CTC). The findings revealed a great potential of CSV in detecting CTC in addition to the conventional EpCAM marker for comprehensive CTC detection.40,41 Furthermore, a combination study on CTC detection using CSV and EpCAM as the marker to detect both epithelial and mesenchymal phenotype of CTC. The study was done to monitor breast cancer therapeutic response. The findings demonstrate the power and reliability of both methods in determining the treatment response of metastatic breast cancer patients.42
As a well-known EMT marker, it is expected to evaluate the expression level of vimentin in cancer studies. Furthermore, exploring some drugs revealed some intriguing findings in how they targeted the rearrangement of vimentin in cancer. One of the active compounds that has been investigated is Withaferin A. Withaferin A is a phytocompound classified as steroidal lactones extracted from the Withania somnifera, known as Ashwagandha, a winter cherry plant found in India. The extract from various parts of this ancient Ayurvedic medicine has been used to treat various medical conditions, including neurological conditions such as Alzheimer’s, to improve memory, enhance neurological conditions, and reduce stress.43–45 The extract of this plant contained various alkaloids, including Withaferin A, which has anticancer properties.43,46,47
In various studies, Withaferin A (WFA) has been discovered to demonstrate cancer toxicity,48 and inhibition of cancer proliferation,49 angiogenesis, migration, invasion, and metastasis,50 especially in breast cancer studies, whether as a single agent or in combination with other active compounds (Table 2). Intriguing findings demonstrated the anti-migration effect of WFA on breast cancer cells. The study showed that WFA suppressed cell migration while not suppressing vimentin expression, as indicated by the insignificant difference in vimentin mRNA expression in the cells treated by WFA. However, microscopic observation showed less distinct formation of lamellipodia and vimentin formation in the leading edge of the cells of WFA-treated cells. This finding suggests that vimentin’s function in forming lamellipodia has been disrupted through the disassembly of vimentin by the interaction of WFA and vimentin proteins.50
 |
Table 2 Studies of Withaferin A and Metformin on Breast Cancer |
Another active compound with intriguing effects on vimentin in cancer cells is metformin. This first-line drug commonly used for Type-2 diabetes patients, metformin, has emerged in cancer studies with its positive impact on cancer. Metformin itself is regarded as the first-line treatment for most type-2 diabetes patients.64 Various findings reported the favorable effect of metformin on cancer therapy, which includes the antiproliferative effect,65,66 anti-migration,67 inhibition of EMT,68 and its potential to assist cancer immunotherapy.69
A study on metformin reveals that it inhibits the migration of endometrial cancer cells through the PI3K/AKT/MDM2 pathway.67 Furthermore, this drug attenuates the EMT process of endometrial cancer by affecting the intermediate filament expression, including the upregulation of Pan-keratin and suppression of vimentin.68 Other studies by Valee demonstrate that metformin suppresses the EMT process in gastric cancer.70 Metformin is also reported to exhibit an inhibition effect on cancer cell stemness and EMT by inhibiting the Wnt3a/β-catenin pathway, by which one of the downstream is the expression of vimentin.71,72 Further studies discovered that metformin specifically targeting vimentin has a comparable suppression effect with vimentin-siRNA.70,73
Nestin
Nestin cytoskeleton is classified as Type IV intermediate filament, initially typically identified in the neuroepithelial progenitor/stem cell in the earlier year.74 Hence, nestin has been known as a biomarker for central nervous system progenitor cells. Despite this, further studies also demonstrate the expression of nestin in other types of progenitor cells, including endothelium, testes, teeth, bone marrow, intestines, muscles, and other tissues.75–81 In-depth studies by Sahlgren et al describe the involvement of nestin in cell regulation, specifically related to its phosphorylation by CDK1 (cdc2 kinase) and CDK5. The rearrangement of nestin was reported to stimulate cell proliferation and survival against oxidative stress-induced cell death (Figure 2).82–84
 |
Figure 2 The role of nestin in breast cancer progression. |
Li et al published the first identification of nestin expression in breast cancer. It was found specifically in the basal layer of highly regenerative cells of normal mammary glands. Nestin expression was colocalized with other myoepithelial or basal epithelial markers, such as p63 and CK14—similar protein profiles in basal-like breast cancer cells, including high nestin, p63, and CK14 expression. In another study, nestin expression is also abundantly found in BRCA1-related breast cancer. Hence, it demonstrates the phenotype of a progenitor-like cell of a BRCA1-related tumor with high aggressiveness and reduced overall survival of the specific breast cancer.85,86
Nestin was also reported to be involved in drug resistance to breast cancer. A study by Feng et al demonstrates the correlation of a long noncoding RNA (lncRNA) ENST869 with nestin expression in breast cancer cells. In the study, the inhibition of LncRNA ENST869 by siRNA3 resulted in the downregulation of nestin expression. Moreover, the decrease of nestin led to the elevation of an anticancer drug, Chidamide, in its effectiveness on breast cancer cells, implying nestin’s role as a crucial drug resistance factor in breast cancer.20
Another study on breast cancer reported that nestin is specifically expressed during the development of the endothelium of blood vessels. Thus, high nestin expression is found in microvessels during angiogenesis. Furthermore, high nestin expression microvessel density (Nestin+MVD) was related to shorter overall survival (OS) in breast cancer patients. Hence, Nestin+MVD is proposed to be an independent prognostic factor in breast cancer.17 A follow-up study further confirmed the importance of nestin as a poor prognostic factor for breast cancer, in which the expression of nestin in both mRNA and protein levels is elevated in more aggressive types of breast cancer cell lines.18
Keratin
Keratin or cytokeratin (CK) is classified into Type I and Type II intermediate filaments of epithelial cells, which assemble into filament structures by heterodimerization and polymerization.6,87 Fifty-four human keratins have been characterized, which consist of 37 human epithelial keratins and 17 hair keratins. Type I epithelial keratins consist of 17 subclasses (CK9-CK28), in which numbers CK11, CK21, and CK22 are not used currently due to some circumstances discovered before. Meanwhile, Type II epithelial keratins are classified into 20 subclasses (CK1-CK8 and CK71-CK80), in which CK6 consists of three variables: CK6a, CK6b, and CK6c.88–90
As a part of the cytoskeleton family, keratins are an integral factor in maintaining the structural stability of epithelial cells. Furthermore, the importance of keratin filament is beyond the cellular level due to its interaction with desmosomes and hemidesmosomes, which provide stability for intercell connection and the attachment of the cells to the basal membrane.91 In cancer studies and clinical settings, keratin pattern determination has been extensively utilized to characterize tumors, including metastasis tumors. This exploitation occurs due to the general preservation of keratin pattern expression in tumor cells mimicking the origin cells it derived from.91,92 Cytokeratin subclasses, which are substantially expressed and used in breast cancer typing, include CK5/CK6, CK7, CK8, CK17, CK18, and CK19 (Figure 3).7,91,93
 |
Figure 3 The role of cytokeratin in breast cancer progression. |
Cytokeratin 5 (CK5) and cytokeratin 6 (CK6) are often identified concurrently using bispecific antibody CK5/CK6 for clinical cancer diagnostic purposes, even though they are not functionally associated.21,88 Both cytokeratins are typically expressed in squamous cell types. The CK5 is mainly localized in the basal layer, while the CK6 is mostly expressed in the suprabasal layers. Hence, the identification of CK5/CK6 is utilized to determine the origin of the carcinoma’s squamous cell type in weakly differentiated carcinomas.21,22 In breast cancer studies, CK5/CK6 is often used as a biomarker to identify the subtype of triple-negative breast cancer (TNBC) with basal-like type.94,95 One of the studies showed that patients with CK5/CK6-positive breast cancer have relatively shorter overall survival compared to patients with negative CK5/CK6. Furthermore, maintenance treatment using capecitabine does not increase the overall survival of the CK5/CK6 positive compared to the CK5/CK6-negative group. Thus, CK5/CK6 can serve as a prognostic biomarker, which identifies the poorly prognosis breast cancer.21,22
Cytokeratin 7 (CK7) expression in normal conditions is related to various normal cellular physiological functions such as proliferation, migration, and EMT during embryogenesis. In the same regard, the high level of CK7 expression is also associated with proliferation, migration, and EMT processes in cancer progression.8 As a prognostic biomarker, the higher expression of CK7 is associated with poorer outcomes for the patient. Nevertheless, studies on the attenuation of CK7 expression, such as using the long non-coding RNA CK7 antisense, revealed a tumor-suppressive effect on breast cancer.8,96 The presence of CK7, along with the absence of CK20, has been employed in identifying various original cancer sites of metastase carcinoma, including breast cancer and several other original sites.97 Due to the low specificity of the CK7+/CK20-profile to identify breast cancer origin of metastases tumors, combination evaluation with other biomarkers as comprehensive diagnostic tools for breast cancer metastases cancer has been studied.23
Cytokeratin 8 (CK8) forms an obligate heterodimer with cytokeratin 18 (CK18). CK8/CK18 is dominantly expressed in the luminal type of breast epithelial cells. Thus, it is often used to identify the luminal type of breast cancer.98 The overexpression of CK8/CK18 heterodimer was reported to induce various unfavorable effects on cancer progression and treatment. In earlier years, it was discovered that overexpression of CK8/CK18 leads to multiple drug chemoresistant in cancer.19,99,100 These cytokeratin dimers also demonstrate the protection effect of breast cancer against apoptosis through the TNF-related apoptosis-inducing ligand pathway.101 CK8/CK18 also acts as a substrate of Akt, stabilizing Akt and possibly assisting cancer progression and metastases. Meanwhile, the phosphorylation of CK18 of the CK8/CK18 heterodimer inhibits Akt-CK8/18 protein interaction.102
Cytokeratin 17 (CK17) can be found in the basal type of epithelial cells, which regulate many physiological processes of the cells, including cell growth and proliferation.103 Breast cancer with CK17-positive is usually associated with increased cancer aggressiveness and chemoresistant or low sensitivity against chemotherapy drugs, especially in the TNBC subtype. Suppression of CK17 leads to increased sensitivity of doxorubicin-resistant breast cancer towards doxorubicin treatment. However, the loss of CK17 also promotes the activation of EMT and the acquirement of mesenchymal cells characteristic of breast cancer.104,105 Interestingly, the opposite role of CK17 is found in HER2HIGH breast cancer, in which the suppression of CK17 expression leads to poor prognosis of the disease.106
Cytokeratin 19 (CK19) is the tiniest size of cytokeratin protein with myriad pattern expressions found in various cells, including muscle cells, epithelial cells, and epithelial stem cells.88,107 The expression of CK19 is essential for cell proliferation and has also been reported to be correlated with poor prognosis in several types of cancer.108,109 Intriguingly, the role of CK19 as a prognosis biomarker in breast cancer is still debatable. The expression of CK19 is needed in luminal breast cancer cells to proliferate and regulate the cell cycle by interacting and stabilizing cyclin D3.110 The expression of CK19 is often evaluated in its soluble state and is utilized to monitor the development of breast cancer.91,93,111 The peripheral blood of a normal person does not contain CK19 mRNA.
Meanwhile, the presence of CK19 mRNA is found in almost 50% of non-metastatic breast cancer and over 50% positive rate for the subjects with metastatic breast cancer. Thus, it is correlated with the poor outcome.112 However, another study reveals a fascinating finding: the expression of CK19 in aggressive breast cancer is lower than in the less aggressive type of breast cancer. Furthermore, the study demonstrates that cancer cells are inhibited in cell proliferation, migration, and chemoresistance.113
Last but not least, there are emerging discussions on the role of cell-surface keratin (CSK) in cancer progression. Previous studies reported that the expression of cell-surface keratin 8 (CSK8) is involved in the mechanism by which cancer cells evade immune surveillance. The presence of CK8/CK18 or CK8/CK19 on the surface of metastatic carcinoma cells concealed the major histocompatibility complex class I (MCH I) molecules, thus avoiding elimination by cytotoxic T-cells due to MHC I – T-cell receptor (TRC) interaction inhibition.3,114,115
Another intriguing cell surface keratin that has been studied is cell-surface keratin 1 (CSK1). This molecule was found to be overexpressed in malignant cells. Previous research on breast cancer developed cytotoxic peptides that specifically targeted CSK1 in breast cancer cells, known as peptide 18–4, the analog of cytotoxic peptide p160. Both p160 and its analog peptide 18–4 could internalize into the cancer cells through the binding with CSK1, while peptide 18–4 does not affect the viability of cancer cells.116 Recent studies by Ziaei et al revealed the development of peptide-drug conjugated, which targets keratin 1 in triple-negative breast cancer cells and the tumor xenograft mice model. The team developed conjugation of peptide 18–4 with doxorubicin (Dox). The peptide 18-4–Dox conjugated demonstrated high cytotoxicity comparable with free Dox on TNBC cells.117 Furthermore, the TNBC xenograft model displayed a greater accumulation of Dox in the peptide-Dox conjugated group in the breast cancer tissue compared to the group treated with free Dox. Meanwhile, the accumulation of Dox in other normal tissues, including the liver, heart, and lungs, is significantly lower for the peptide-Dox group compared to the free Dox group. Thus, these findings imply the potential of CSK1 as a receptor for targeted therapy of TNBC.118,119
Conclusion
In cancer studies, the role of intermediate filament, specifically vimentin, nestin, and keratin, is often mentioned as various biomarkers, whether diagnostic or prognostic. Various intermediate filaments are frequently cited to be related to the progression of cancer. Besides its role in determining the appropriate treatment procedure for the patients, the role of these intermediate filaments as target therapy is also an intriguing topic to explore. The overexpression of vimentin and nestin generally indicates a poor prognosis of cancer. However, due to the myriad subclass of keratin and evidence showing high or low expression of keratin in breast cancer, no conclusion can be drawn for keratin expression in general. In some cases, some keratin subclasses were found to be overexpressed in breast cancer, including CK5/CK6, CK7, CK8/CK18, and CK19, which are involved in angiogenesis promotion, metastasis, and chemoresistance.98,102,105,112,120 On the other hand, some studies also reported a protective effect of keratin 17 against cancer.106 Furthermore, other studies on keratin, such as CK19 were found to be downregulated in highly aggressive breast cancer compared to less aggressive cancers.113
The range of promising anticancer drug research has increased, encompassing natural-derived bioactive compounds and chemical synthesis products. The exploration of potential anticancer drugs even reaches the exploration of other medications used to treat other ailments. Most potential substances were evaluated for their cytotoxicity, antiproliferative, and anti-metastasis effects on cancer cells. Some studies on active compounds, such as Withaferin A and Metformin, demonstrated the substance’s impact on vimentin in cancer cells concerning the cancer progression and anti-metastasis effect.43,46,49,65,69,73,121 In addition, exploration in peptide-drug conjugation targeting cell-surface keratin 1 displayed the potential of cell-surface keratin as a receptor, especially for TNBC therapy development.116,117,119 However, the effect of active compounds on other intermediate filaments, such as nestin, is still underexplored.
Acknowledgments
We want to thank the Faculty of Medicine, Universitas Padjadjaran, for their substantial support in completing this study.
Author Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, data analysis and interpretation, or all these areas; took part in drafting, revising, or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Disclosure
The authors report no conflicts of interest in this work.
References
1. Kalluri R, Weinberg RA. The basics of epithelial-mesenchymal transition. J Clin Invest. 2009;119(6):1420–1428. doi:10.1172/JCI39104
2. Amack JD. Cellular dynamics of EMT: lessons from live in vivo imaging of embryonic development. Cell Commun Signal. 2021;19(1):79. doi:10.1186/s12964-021-00761-8
3. Ogunnigbagbe O, Bunick CG, Kaur K. Keratin 1 as a cell-surface receptor in cancer. Biochim Biophys Acta Rev Cancer. 2022;1877(1):188664. doi:10.1016/j.bbcan.2021.188664
4. Zeng Y, Du W, Huang Z, et al. Hsa_circ_0060467 promotes breast cancer liver metastasis by complexing with eIF4A3 and sponging miR-1205. Cell Death Discov. 2023;9(1):153. doi:10.1038/s41420-023-01448-4
5. Liu P, Wang Z, Ou X, et al. The FUS/circEZH2/KLF5/ feedback loop contributes to CXCR4-induced liver metastasis of breast cancer by enhancing epithelial-mesenchymal transition. Mol Cancer. 2022;21(1):198. doi:10.1186/s12943-022-01653-2
6. Sharma P, Alsharif S, Fallatah A, Chung BM. Intermediate filaments as effectors of cancer development and metastasis: a focus on keratins, vimentin, and nestin. Cells. 2019;8(5). doi:10.3390/cells8050497
7. Benacka R, Szaboova D, Gulasova Z, Hertelyova Z, Radonak J. Classic and new markers in diagnostics and classification of breast cancer. Cancers. 2022;14(21). doi:10.3390/cancers14215444
8. Hosseinalizadeh H, Hussain QM, Poshtchaman Z, et al. Emerging insights into keratin 7 roles in tumor progression and metastasis of cancers. Front Oncol. 2023;13:1243871. doi:10.3389/fonc.2023.1243871
9. ElFeky AW, Saied EM, Shawky H, Sadaka E. Prognostic value of basal markers (Epidermal Growth Factor Receptor ”EGFR” And Cytokeratin 5/6) expression in triple-negative invasive breast cancer. J Pak Med Assoc. 2023;73(4):S161–S166. doi:10.47391/JPMA.EGY-S4-33
10. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48. doi:10.3322/caac.21763
11. Sung H, Ferlay J, Siegel RL, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–249. doi:10.3322/caac.21660
12. Omary MB. ”IF-pathies”: a broad spectrum of intermediate filament-associated diseases. J Clin Invest. 2009;119(7):1756–1762. doi:10.1172/JCI39894
13. Alberts B, Hopkin K, Johnson A, et al. Cytoskeleton. In: Twitchell B, Morales M, editors. Essential Cell Biology.
14. Institute NC. Cancer staging. 2024. Available from: https://www.cancer.gov/about-cancer/diagnosis-staging/staging.
15. Castaneda M, den Hollander P, Kuburich NA, Rosen JM, Mani SA. Mechanisms of cancer metastasis. Semin Cancer Biol. 2022;87:17–31. doi:10.1016/j.semcancer.2022.10.006
16. Lamouille S, Xu J, Derynck R. Molecular mechanisms of epithelial-mesenchymal transition. Nat Rev Mol Cell Biol. 2014;15(3):178–196. doi:10.1038/nrm3758
17. Nowak A, Grzegrzolka J, Paprocka M, et al. Nestin-positive microvessel density is an independent prognostic factor in breast cancer. Int J Oncol. 2017;51(2):668–676. doi:10.3892/ijo.2017.4057
18. Nowak A, Grzegrzolka J, Kmiecik A, Piotrowska A, Matkowski R, Dziegiel P. Role of nestin expression in angiogenesis and breast cancer progression. Int J Oncol. 2018;52(2):527–535. doi:10.3892/ijo.2017.4223
19. Anderson JM, Heindl LM, Bauman PA, Ludi CW, Dalton WS, Cress AE. Cytokeratin expression results in a drug-resistant phenotype to six different chemotherapeutic agents. Clin Cancer Res. 1996;2(1):97–105.
20. Feng X, Han H, Guo Y, Feng X, Guo S, Zhou W. LncRNA ENST869 targeting nestin transcriptional region to affect the pharmacological effects of chidamide in breast cancer cells. Front Oncol. 2022;12:874343. doi:10.3389/fonc.2022.874343
21. Kaufmann O, Fietze E, Mengs J, Dietel M. Value of p63 and cytokeratin 5/6 as immunohistochemical markers for the differential diagnosis of poorly differentiated and undifferentiated carcinomas. Am J Clin Pathol. 2001;116(6):823–830. doi:10.1309/21TW-2NDG-JRK4-PFJX
22. Volkel C, De Wispelaere N, Weidemann S, et al. Cytokeratin 5 and cytokeratin 6 expressions are unconnected in normal and cancerous tissues and have separate diagnostic implications. Virchows Arch. 2022;480(2):433–447. doi:10.1007/s00428-021-03204-4
23. Statz E, Jorns JM. Cytokeratin 7, GATA3, and SOX-10 is a comprehensive panel in diagnosing triple negative breast cancer brain metastases. Int J Surg Pathol. 2021;29(5):470–474. doi:10.1177/1066896921990717
24. Satala CB, Jung I, Bara TJ, et al. Primary rhabdomyosarcoma: an extremely rare and aggressive variant of male breast cancer. World J Clin Cases. 2020;8(19):4466–4474. doi:10.12998/wjcc.v8.i19.4466
25. Dias P, Kumar P, Marsden HB, et al. Evaluation of desmin as a diagnostic and prognostic marker of childhood rhabdomyosarcomas and embryonal sarcomas. Br J Cancer. 1987;56:361–365.
26. Alberts B, Hopkin K, Johnson A, et al. Essential Cell Biology.
27. Strouhalova K, Prechova M, Gandalovicova A, Brabek J, Gregor M, Rosel D. Vimentin intermediate filaments as potential target for cancer treatment. Cancers. 2020;12(1). doi:10.3390/cancers12010184
28. Stankevicins LDC, Urbanska M, Flormann DA, et al. Vimentin provides the mechanical resilience required for amoeboid migration and protection of the nucleus. bioRxiv. 2019. doi:10.1101/720946
29. Danielsson F, Peterson MK, Caldeira Araujo H, Lautenschlager F, Gad AKB. Vimentin diversity in health and disease. Cells. 2018;7(10). doi:10.3390/cells7100147
30. Liu CY, Lin HH, Tang MJ, Wang YK. Vimentin contributes to epithelial-mesenchymal transition cancer cell mechanics by mediating cytoskeletal organization and focal adhesion maturation. Oncotarget. 2015;6(18):15966–15983. doi:10.18632/oncotarget.3862
31. Leube RE, Moch M, Windoffer R. Intermediate filaments and the regulation of focal adhesion. Curr Opin Cell Biol. 2015;32:13–20. doi:10.1016/j.ceb.2014.09.011
32. Gregor M, Osmanagic-Myers S, Burgstaller G, et al. Mechanosensing through focal adhesion-anchored intermediate filaments. FASEB J. 2014;28(2):715–729. doi:10.1096/fj.13-231829
33. Winter M, Meignan S, Volkel P, et al. Vimentin promotes the aggressiveness of triple negative breast cancer cells surviving chemotherapeutic treatment. Cells. 2021;10(6). doi:10.3390/cells10061504
34. Zhu QS, Rosenblatt K, Huang KL, et al. Vimentin is a novel AKT1 target mediating motility and invasion. Oncogene. 2011;30(4):457–470. doi:10.1038/onc.2010.421
35. Ivaska J. Vimentin: central hub in EMT induction? Small GTPases. 2011;2(1):51–53. doi:10.4161/sgtp.2.1.15114
36. Milovanovic J, Todorovic-Rakovic N, Abu Rabi Z. The role of interleukin 8 and matrix metalloproteinases 2 and 9 in breast cancer treated with tamoxifen. J BUON. 2017;22(3):628–637.
37. Li B, Lou G, Zhou J. MT1‑MMP promotes the proliferation and invasion of gastric carcinoma cells via regulating vimentin and E‑cadherin. Mol Med Rep Apr. 2019;19(4):2519–2526. doi:10.3892/mmr.2019.9918
38. Sadeghi N, Fazli G, Bayat AA, et al. Cell surface vimentin detection in cancer cells by peptide-based monoclonal antibody. Avicenna J Med Biotechnol. 2023;15(2):68–75.
39. Batth IS, Li S. Discovery of Cell-Surface Vimentin (CSV) as a sarcoma target and development of CSV-Targeted IL12 immune therapy. Adv Exp Med Biol. 2020;1257:169–178. doi:10.1007/978-3-030-43032-0_14
40. Li H, Zhu YZ, Xu L, et al. Exploring new frontiers: cell surface vimentin as an emerging marker for circulating tumor cells and a promising therapeutic target in advanced gastric cancer. J Exp Clin Cancer Res. 2024;43(1):129. doi:10.1186/s13046-024-03043-6
41. Yu J, Yang M, Peng T, Liu Y, Cao Y. Evaluation of cell surface vimentin positive circulating tumor cells as a prognostic biomarker for stage III/IV colorectal cancer. Sci Rep. 2023;13(1):18791. doi:10.1038/s41598-023-45951-1
42. Satelli A, Brownlee Z, Mitra A, Meng QH, Li S. Circulating tumor cell enumeration with a combination of epithelial cell adhesion molecule- and cell-surface vimentin-based methods for monitoring breast cancer therapeutic response. Clin Chem. 2015;61(1):259–266. doi:10.1373/clinchem.2014.228122
43. Sultana T, Okla MK, Ahmed M, et al. Withaferin A: from ancient remedy to potential drug candidate. Molecules. 2021;26(24). doi:10.3390/molecules26247696
44. Ram N, Peak SL, Perez AR, Jinwal UK. Implications of withaferin A in neurological disorders. Neural Regen Res. 2021;16(2):304–305. doi:10.4103/1673-5374.290894
45. Das R, Rauf A, Akhter S, et al. Role of withaferin A and its derivatives in the management of alzheimer’s disease: recent trends and future perspectives. Molecules. 2021;26(12). doi:10.3390/molecules26123696
46. Xing Z, Su A, Mi L, et al. Withaferin A: a dietary supplement with promising potential as an anti-tumor therapeutic for cancer treatment - pharmacology and mechanisms. Drug Des Devel Ther. 2023;17:2909–2929. doi:10.2147/DDDT.S422512
47. Straughn AR, Kakar SS. Withaferin A: a potential therapeutic agent against COVID-19 infection. J Ovarian Res. 2022;13(1):79. doi:10.1186/s13048-020-00684-x
48. Sehrawat A, Samanta SK, Hahm ER, St Croix C, Watkins S, Singh SV. Withaferin A-mediated apoptosis in breast cancer cells is associated with alterations in mitochondrial dynamics. Mitochondrion. 2019;47:282–293. doi:10.1016/j.mito.2019.01.003
49. Zuniga R, Concha G, Cayo A, et al. Withaferin A suppresses breast cancer cell proliferation by inhibition of the two-pore domain potassium (K2P9) channel TASK-3. Biomed Pharmacother. 2020;129:110383. doi:10.1016/j.biopha.2020.110383
50. Thaiparambil JT, Bender L, Ganesh T, et al. Withaferin A inhibits breast cancer invasion and metastasis at sub-cytotoxic doses by inducing vimentin disassembly and serine 56 phosphorylation. Int J Cancer. 2011;129(11):2744–2755. doi:10.1002/ijc.25938
51. Sari AN, Dhanjal JK, Elwakeel A, et al. A low dose combination of withaferin A and caffeic acid phenethyl ester possesses anti-metastatic potential in vitro: molecular targets and mechanisms. Cancers. 2022;14(3). doi:10.3390/cancers14030787
52. Lee J, Hahm ER, Marcus AI, Singh SV. Withaferin A inhibits experimental epithelial-mesenchymal transition in MCF-10A cells and suppresses vimentin protein level in vivo in breast tumors. Mol Carcinog. 2015;54(6):417–429. doi:10.1002/mc.22110
53. Rahman MM, Wu H, Tollefsbol TO. A novel combinatorial approach using sulforaphane- and withaferin A-rich extracts for prevention of estrogen receptor-negative breast cancer through epigenetic and gut microbial mechanisms. Sci Rep. 2024;14(1):12091. doi:10.1038/s41598-024-62084-1
54. Samanta SK, Sehrawat A, Kim SH, et al. Disease subtype-independent biomarkers of breast cancer chemoprevention by the ayurvedic medicine phytochemical withaferin A. J Natl Cancer Inst. 2017;109(6). doi:10.1093/jnci/djw293
55. Meghalatha TS, Muninathan N. Antitumor activity of withaferin-A and propolis in benz (a) pyrene-induced breast cancer. Bioinformation. 2022;18(9):841–844. doi:10.6026/97320630018841
56. Kawakita E, Yang F, Kumagai A, et al. Metformin mitigates DPP-4 inhibitor-induced breast cancer metastasis via suppression of mTOR signaling. Mol Cancer Res. 2021;19(1):61–73. doi:10.1158/1541-7786.MCR-20-0115
57. Cingir Koker S, Yalcin B, Dogan Turacli I. Metformin resistant MDA-MB-468 cells exhibit EMT-like phenotype and increased migration capacity. Mol Biol Rep. 2022;49(7):5973–5984. doi:10.1007/s11033-022-07381-6
58. Tabari AR, Gavidel P, Sabouni F, Gardaneh M. Synergy between sublethal doses of shikonin and metformin fully inhibits breast cancer cell migration and reverses epithelial-mesenchymal transition. Mol Biol Rep. 2022;49(6):4307–4319. doi:10.1007/s11033-022-07265-9
59. Jang SY, Kim A, Kim JK, et al. Metformin inhibits tumor cell migration via down-regulation of MMP9 in tamoxifen-resistant breast cancer cells. Anticancer Res. 2014;34(8):4127–4134.
60. Esparza-Lopez J, Alvarado-Munoz JF, Escobar-Arriaga E, Ulloa-Aguirre A, de Jesus Ibarra-Sanchez M. Metformin reverses mesenchymal phenotype of primary breast cancer cells through STAT3/NF-kappaB pathways. BMC Cancer. 2019;19(1):728. doi:10.1186/s12885-019-5945-1
61. Schexnayder C, Broussard K, Onuaguluchi D, et al. Metformin inhibits migration and invasion by suppressing ROS production and COX2 expression in MDA-MB-231 breast cancer cells. Int J Mol Sci. 2018;19(11). doi:10.3390/ijms19113692
62. Song J, Du J, Han L, Lin X, Fan C, Chen G. The effect of metformin on triple-negative breast cancer cells and nude mice. Altern Ther Health Med. 2023;29(8):389–395.
63. Wang JC, Li GY, Wang B, et al. Metformin inhibits metastatic breast cancer progression and improves chemosensitivity by inducing vessel normalization via PDGF-B downregulation. J Exp Clin Cancer Res. 2019;38(1):235. doi:10.1186/s13046-019-1211-2
64. Triggle CR, Mohammed I, Bshesh K, et al. Metformin: is it a drug for all reasons and diseases? Metabolism. 2022;133:155223. doi:10.1016/j.metabol.2022.155223
65. Cejuela M, Martin-Castillo B, Menendez JA, Pernas S. Metformin and breast cancer: where are we now? Int J Mol Sci. 2022;23(5). doi:10.3390/ijms23052705
66. Sharma A, Bandyopadhayaya S, Chowdhury K, et al. Metformin exhibited anticancer activity by lowering cellular cholesterol content in breast cancer cells. PLoS One. 2019;14(1):e0209435. doi:10.1371/journal.pone.0209435
67. Qiang P, Shao Y, Sun YP, Zhang J, Chen LJ. Metformin inhibits proliferation and migration of endometrial cancer cells through regulating PI3K/AKT/MDM2 pathway. Eur Rev Med Pharmacol Sci. 2019;23(4):1778–1785. doi:10.26355/eurrev_201902_17140
68. Laskov I, Abou-Nader P, Amin O, et al. Metformin increases E-cadherin in tumors of diabetic patients with endometrial cancer and suppresses epithelial-mesenchymal transition in endometrial cancer cell lines. Int J Gynecol Cancer. 2016;26(7):1213–1221. doi:10.1097/IGC.0000000000000761
69. Panaampon J, Zhou Y, Saengboonmee C. Metformin as a booster of cancer immunotherapy. Int Immunopharmacol. 2023;121:110528. doi:10.1016/j.intimp.2023.110528
70. Valaee S, Yaghoobi MM, Shamsara M. Metformin inhibits gastric cancer cells metastatic traits through suppression of epithelial-mesenchymal transition in a glucose-independent manner. PLoS One. 2017;12(3):e0174486. doi:10.1371/journal.pone.0174486
71. Zhang C, Wang Y. Metformin attenuates cells stemness and epithelial‑mesenchymal transition in colorectal cancer cells by inhibiting the Wnt3a/beta‑catenin pathway. Mol Med Rep. 2019;19(2):1203–1209. doi:10.3892/mmr.2018.9765
72. Dongre A, Weinberg RA. New insights into the mechanisms of epithelial-mesenchymal transition and implications for cancer. Nat Rev Mol Cell Biol. 2019;20(2):69–84. doi:10.1038/s41580-018-0080-4
73. Valaee S, Shamsara M, Yaghoobi MM. Metformin is a novel suppressor for vimentin in human gastric cancer cell line. Int J Mol Cell Med. 2021;10(3):200–206. doi:10.22088/IJMCM.BUMS.10.3.200
74. Lendahl U, Zimmerman LB, McKay RD. CNS stem cells express a new class of intermediate filament protein. Cell. 1990;60(4):585–595. doi:10.1016/0092-8674(90)90662-x
75. Xie L, Zeng X, Hu J, Chen Q. Characterization of nestin, a selective marker for bone marrow derived mesenchymal stem cells. Stem Cells Int. 2015;2015:762098. doi:10.1155/2015/762098
76. Vanderwinden JM, Gillard K, De Laet MH, Messam CA, Schiffmann SN. Distribution of the intermediate filament nestin in the muscularis propria of the human gastrointestinal tract. Cell Tissue Res. 2002;309(2):261–268. doi:10.1007/s00441-002-0590-3
77. Kachinsky AM, Dominov JA, Miller JB. Myogenesis and the intermediate filament protein, nestin. Dev Biol. 1994;165(1):216–228. doi:10.1006/dbio.1994.1248
78. About I, Laurent-Maquin D, Lendahl U, Mitsiadis TA. Nestin expression in embryonic and adult human teeth under normal and pathological conditions. Am J Pathol. 2000;157(1):287–295. doi:10.1016/S0002-9440(10)64539-7
79. Lobo MV, Arenas MI, Alonso FJ, et al. Nestin, a neuroectodermal stem cell marker molecule, is expressed in Leydig cells of the human testis and in some specific cell types from human testicular tumours. Cell Tissue Res. 2004;316(3):369–376. doi:10.1007/s00441-003-0848-4
80. Suzuki S, Namiki J, Shibata S, Mastuzaki Y, Okano H. The neural stem/progenitor cell marker nestin is expressed in proliferative endothelial cells, but not in mature vasculature. J Histochem Cytochem. 2010;58(8):721–730. doi:10.1369/jhc.2010.955609
81. Sejersen T, Lendahl U. Transient expression of the intermediate filament nestin during skeletal muscle development. J Cell Sci. 1993;106(Pt 4):1291–1300. doi:10.1242/jcs.106.4.1291
82. Sahlgren CM, Pallari HM, He T, Chou YH, Goldman RD, Eriksson JE. A nestin scaffold links Cdk5/p35 signaling to oxidant-induced cell death. EMBO J. 2006;25(20):4808–4819. doi:10.1038/sj.emboj.7601366
83. Sahlgren CM, Mikhailov A, Hellman J, et al. Mitotic reorganization of the intermediate filament protein nestin involves phosphorylation by cdc2 kinase. J Biol Chem. 2001;276(19):16456–16463. doi:10.1074/jbc.M009669200
84. Sahlgren CM, Mikhailov A, Vaittinen S, et al. Cdk5 regulates the organization of Nestin and its association with p35. Mol Cell Biol. 2003;23(14):5090–5106. doi:10.1128/MCB.23.14.5090-5106.2003
85. Li H, Cherukuri P, Li N, et al. Nestin is expressed in the basal/myoepithelial layer of the mammary gland and is a selective marker of basal epithelial breast tumors. Cancer Res. 2007;67(2):501–510. doi:10.1158/0008-5472.CAN-05-4571
86. Kruger K, Wik E, Knutsvik G, et al. Expression of Nestin associates with BRCA1 mutations, a basal-like phenotype and aggressive breast cancer. Sci Rep. 2017;7(1):1089. doi:10.1038/s41598-017-00862-w
87. Ho M, Thompson B, Fisk JN, et al. Update of the keratin gene family: evolution, tissue-specific expression patterns, and relevance to clinical disorders. Hum Genomics. 2022;16(1):1. doi:10.1186/s40246-021-00374-9
88. Moll R, Franke WW, Schiller DL, Geiger B, Krepler R. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell. 1982;31(1):11–24. doi:10.1016/0092-8674(82)90400-7
89. Zhang N, Zhang R, Zou K, et al. Keratin 23 promotes telomerase reverse transcriptase expression and human colorectal cancer growth. Cell Death Dis. 2017;8(7):e2961. doi:10.1038/cddis.2017.339
90. Schweizer J, Bowden PE, Coulombe PA, et al. New consensus nomenclature for mammalian keratins. J Cell Biol. 2006;174(2):169–174. doi:10.1083/jcb.200603161
91. Moll R, Divo M, Langbein L. The human keratins: biology and pathology. Histochem Cell Biol. 2008;129(6):705–733. doi:10.1007/s00418-008-0435-6
92. Effenberger KE, Borgen E, Eulenburg CZ, et al. Detection and clinical relevance of early disseminated breast cancer cells depend on their cytokeratin expression pattern. Breast Cancer Res Treat. 2011;125(3):729–738. doi:10.1007/s10549-010-0911-2
93. Marrakchi R, Ouerhani S, Benammar S, et al. Detection of cytokeratin 19 mRNA and CYFRA 21-1 (cytokeratin 19 fragments) in blood of Tunisian women with breast cancer. Int J Biol Markers. 2008;23(4):238–243. doi:10.1177/172460080802300407
94. Nielsen TO, Hsu FD, Jensen K, et al. Immunohistochemical and clinical characterization of the basal-like subtype of invasive breast carcinoma. Clin Cancer Res. 2004;10(16):5367–5374. doi:10.1158/1078-0432.CCR-04-0220
95. Abd El-Rehim DM, Pinder SE, Paish CE, et al. Expression of luminal and basal cytokeratins in human breast carcinoma. J Pathol. 2004;203(2):661–671. doi:10.1002/path.1559
96. Zhao Z, Meng M, Yao J, et al. The long non-coding RNA keratin-7 antisense acts as a new tumor suppressor to inhibit tumorigenesis and enhance apoptosis in lung and breast cancers. Cell Death Dis. 2023;14(4):293. doi:10.1038/s41419-023-05802-3
97. Selves J, Long-Mira E, Mathieu MC, Rochaix P, Ilie M. Immunohistochemistry for diagnosis of metastatic carcinomas of unknown primary site. Cancers. 2018;10(4). doi:10.3390/cancers10040108
98. Hashmi AA, Naz S, Hashmi SK, et al. Cytokeratin 5/6 and cytokeratin 8/18 expression in triple negative breast cancers: clinicopathologic significance in South-Asian population. BMC Res Notes. 2018;11(1):372. doi:10.1186/s13104-018-3477-4
99. Bauman PA, Dalton WS, Anderson JM, Cress AE. Expression of cytokeratin confers multiple drug resistance. Proc Natl Acad Sci. 1994;91(12):5311–5314. doi:10.1073/pnas.91.12.5311
100. Zhang B, Wang J, Liu W, et al. Cytokeratin 18 knockdown decreases cell migration and increases chemosensitivity in non-small cell lung cancer. J Cancer Res Clin Oncol. 2016;142(12):2479–2487. doi:10.1007/s00432-016-2253-x
101. Bozza WP, Zhang Y, Zhang B. Cytokeratin 8/18 protects breast cancer cell lines from TRAIL-induced apoptosis. Oncotarget. 2018;9(33):23264–23273. doi:10.18632/oncotarget.25297
102. Lim Y, Kim S, Yoon HN, Ku NO. Keratin 8/18 regulate the Akt signaling pathway. Int J Mol Sci. 2021;22(17). doi:10.3390/ijms22179227
103. Yang L, Zhang S, Wang G. Keratin 17 in disease pathogenesis: from cancer to dermatoses. J Pathol. 2019;247(2):158–165. doi:10.1002/path.5178
104. McGinn O, Riley D, Finlay-Schultz J, Paul KV, Kabos P, Sartorius CA. Cytokeratins 5 and 17 maintain an aggressive epithelial state in basal-like breast cancer. Mol Cancer Res. 2022;20(9):1443–1455. doi:10.1158/1541-7786.MCR-21-0866
105. Wu L, Ding W, Wang X, Li X, Yang J. Interference KRT17 reverses doxorubicin resistance in triple-negative breast cancer cells by Wnt/beta-catenin signaling pathway. Genes Genomics. 2023;45(10):1329–1338. doi:10.1007/s13258-023-01437-y
106. Tang S, Liu W, Yong L, et al. Reduced expression of KRT17 predicts poor prognosis in HER2(high) breast cancer. Biomolecules. 2022;12(9). doi:10.3390/biom12091183
107. Petersen OW, Polyak K. Stem cells in the human breast. Cold Spring Harb Perspect Biol. 2010;2(5):a003160. doi:10.1101/cshperspect.a003160
108. Kabir NN, Ronnstrand L, Kazi JU. Keratin 19 expression correlates with poor prognosis in breast cancer. Mol Biol Rep. 2014;41(12):7729–7735. doi:10.1007/s11033-014-3684-6
109. Kim H, Choi GH, Na DC, et al. Human hepatocellular carcinomas with ”Stemness”-related marker expression: keratin 19 expression and a poor prognosis. Hepatology. 2011;54(5):1707–1717. doi:10.1002/hep.24559
110. Sharma P, Alsharif S, Bursch K, et al. Keratin 19 regulates cell cycle pathway and sensitivity of breast cancer cells to CDK inhibitors. Sci Rep. 2019;9(1):14650. doi:10.1038/s41598-019-51195-9
111. Liscia DS, Detoma P, Zanchetta M, et al. The use of CYFRA 21-1 for the detection of breast cancer axillary lymph node metastases in needle washouts of fine-needle aspiration biopsies. Appl Immunohistochem Mol Morphol. 2017;25(3):190–195. doi:10.1097/PAI.0000000000000287
112. Park HS, Han HJ, Lee S, et al. Detection of circulating tumor cells in breast cancer patients using cytokeratin-19 real-time RT-PCR. Yonsei Med J. 2017;58(1):19–26. doi:10.3349/ymj.2017.58.1.19
113. Saha SK, Kim K, Yang G-M, Choi HY, Cho S-G. Cytokeratin 19 (KRT19) has a role in the reprogramming of cancer stem cell-like cells to less aggressive and more drug-sensitive cells. Int J Mol Sci. 2018;19(5):1423. doi:10.3390/ijms19051423
114. Mohme M, Riethdorf S, Pantel K. Circulating and disseminated tumour cells - mechanisms of immune surveillance and escape. Nat Rev Clin Oncol. 2017;14(3):155–167. doi:10.1038/nrclinonc.2016.144
115. Wu MS, Li CH, Ruppert JG, Chang CC. Cytokeratin 8-MHC class I interactions: a potential novel immune escape phenotype by a lymph node metastatic carcinoma cell line. Biochem Biophys Res Commun. 2013;441(3):618–623. doi:10.1016/j.bbrc.2013.10.105
116. Soudy R, Etayash H, Bahadorani K, Lavasanifar A, Kaur K. Breast cancer targeting peptide binds keratin 1: a new molecular marker for targeted drug delivery to breast cancer. Mol Pharm. 2017;14(3):593–604. doi:10.1021/acs.molpharmaceut.6b00652
117. Ziaei E, Saghaeidehkordi A, Dill C, Maslennikov I, Chen S, Kaur K. Targeting triple negative breast cancer cells with novel cytotoxic peptide-doxorubicin conjugates. Bioconjug Chem. 2019;30(12):3098–3106. doi:10.1021/acs.bioconjchem.9b00755
118. Saghaeidehkordi A, Chen S, Yang S, Kaur K. Evaluation of a keratin 1 targeting peptide-doxorubicin conjugate in a mouse model of triple-negative breast cancer. Pharmaceutics. 2021;13(5). doi:10.3390/pharmaceutics13050661
119. Ziaei E, Paiva IM, Yao SJ, et al. Peptide-drug conjugate targeting keratin 1 inhibits triple-negative breast cancer in Mice. Mol Pharm. 2023;20(7):3570–3577. doi:10.1021/acs.molpharmaceut.3c00189
120. Dum D, Menz A, Volkel C, et al. Cytokeratin 7 and cytokeratin 20 expression in cancer: a tissue microarray study on 15,424 cancers. Exp Mol Pathol. 2022;126:104762. doi:10.1016/j.yexmp.2022.104762
121. Suman S, Das TP, Sirimulla S, Alatassi H, Ankem MK, Damodaran C. Withaferin-A suppress AKT induced tumor growth in colorectal cancer cells. Oncotarget. 2016;7(12):13854–13864. doi:10.18632/oncotarget.7351
 © 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.
© 2024 The Author(s). This work is published and licensed by Dove Medical Press Limited. The
full terms of this license are available at https://www.dovepress.com/terms
and incorporate the Creative Commons Attribution
- Non Commercial (unported, 3.0) License.
By accessing the work you hereby accept the Terms. Non-commercial uses of the work are permitted
without any further permission from Dove Medical Press Limited, provided the work is properly
attributed. For permission for commercial use of this work, please see paragraphs 4.2 and 5 of our Terms.